Production of Taxol by Endophytic Fungi Isolated from Roots of Himalayan Yew (Taxus wallichiana Zucc.)
Priyanka Adhikari1, Mithilesh Singh1 and Anita Pandey1, 2,*
1G.B. Pant National Institute of Himalayan Environment, Kosi-Katarmal, Almora-263643, Uttarakhand, India
2Department of Biotechnology, Graphic Era Deemed to be University, Bell Road, Clement Town, Dehradun-248002, Uttarakhand, India
E-mail: anitapandey333@gmail.com; anita.pandey@geu.ac.in
*Corresponding Author
Received 27 April 2022; Accepted 18 August 2022; Publication 06 October 2022
Abstract
Taxol (generic name – Paclitaxel), the most promising chemotherapeutic agent was isolated from bark of different Taxus sp. As Taxus species are threatened with extinction (endangered in Himalaya), thus it is imperative to develop alternate and sustainable method for commercialization and scale up production of paclitaxel. In this respect, physical and chemical parameters are effective and important key points for active compound production particularly by using endophytic microbes. In the present study, five endophytic fungi isolated from the roots of Taxus wallichiana, are tested for paclitaxel production using biochemical and molecular methods. Subsequently, effect of physico-chemical parameters like temperature, pH, incubation time, and medium constituents i.e., salt concentration, carbon and nitrogen sources on paclitaxel production were also analyzed. Among isolates, two of the fungi viz. GBPI_TWR F1 (Penicillium sp.) and GBPI_TWR F5 (Aspergillus sp.) were found to be paclitaxel producing. The genomic DNA samples were sequenced to confirm the presence of two genes viz. 10-deacetylbaccatin III-10-O-acetyl transferase (DBAT) and C-13 phenylpropanoid side chain-CoA acyltransferase (BAPT), implicated in paclitaxel biosynthesis. Both the endophytes showed the amplicons of DBAT and BAPT genes. Results revealed that after optimization of medium components and physical condition, paclitaxel production was increased in both the endophytes, maximum paclitaxel production i.e., 5.45 0.98 mg/L was obtained by GBPI_TWR F5 (Aspergillus sp.) following 10 days of incubation at 15C in optimized S7 liquid medium composition.
Keywords: Taxus wallichiana, endophytes, taxol, Aspergillus, Penicillium.
1 Introduction
Taxol (generic name- Paclitaxel) is a diterpenoid alkaloid that was isolated for the first time from the bark of Taxus brevifolia Nutt. (Wani et al., 1971). Later, many taxol producing Taxus species viz. T. baccata, T. chinensis, T. canadensis, T. wallichiana were identified (Poupat et al., 2000). Among the secondary metabolites with anti-cancer activity, taxol is chemically and pharmaceutically most valuable drug used in treatment of a variety of cancers including refractory ovarian cancer, breast cancer, non-small cell lung cancer, and AIDS related Kaposi’s sarcoma, head and neck carcinoma and other types of cancer (Barbuti et al., 2015).
At present, taxol is extracted from the bark of different Taxus species which grow only in particular geographical areas. Moreover, this gymnosperm genus is very slow growing and almost all Taxus species are under IUCN threatened category (Thomas and Farjon, 2000). Large amount of bark extraction further threats the existence of the plants. To meet the increasing demand of this blockbuster drug and to make it available for poor people, there is need of an alternate, cost effective and sustainable method of its production. Plant tissue culture of Taxus species has brought some hope for its year-round and consistent production. But low yield and plant cell shear resistance have further limited industrial scale production of paclitaxel by this method (Kusari et al., 2014). As the taxol content in Taxus species is very low (0.01–0.03% dry weight), bark of many plants is required for obtaining the required taxol content and lots of yews are demolished in the process (Zaiyou et al., 2015). So, this is not reasonable from the ecological opinion and view. Therefore, it is a necessity to aspect for alternative sources of taxol manufacture to resolve its source. In this view, intensive research efforts are required to develop an efficient and sustainable method for production of this valuable drug without exploiting field grown plants (Somjaipeng et al., 2016; El-Bialy and El-Bastawisy 2020). In this context, microbial fermentation technology offers an efficient and promising alternate approach for taxol production at industrial scale (Kumar et al., 2019).
Following isolation and identification of the first taxol producing fungus (Taxomyces andreanae), worldwide research efforts have been on scaling up taxol production through microbial fermentation (Stierle et al., 1993; Kumar et al., 2019; Abdel-Fatah et al., 2021). Though, so far, taxol production at industrial scale using microbes could not be practically possible owing to low reproducible yield. This suggests the need for further in-depth research on isolation and identification of other potential source of taxol. As it is well established that taxol content in plant depends on geographical location, environmental conditions, and other biotic and abiotic factors (Kusari et al., 2014). It is imperative to screen potential microbes from Taxus spp. of different geographical locations.
Taxus wallichiana Zucc. (English name: Himalayan Yew; Hindi name: Thuner; family Taxaceae) is recognized as a medicinally important evergreen tree that grows under temperate locations of Indian Himalayan region at an altitude range of 1800–3300 m. It is medium sized, slow growing and dioecious gymnosperm. The species has received considerable attention on account of its existing exploitation for the extraction of taxol (Nadeem et al., 2002). To the best of our knowledge, so far, roots of T. wallichiana plant (Himalayan Yew) have never been investigated for paclitaxel producing endophytes. In view of this, the present study aimed to (i) investigate roots of Taxus wallichiana (Himalayan Yew) for paclitaxel producing endophytic fungi and; (ii) optimization of fermentation media (S7) composition for scaling up the paclitaxel production in vitro.
2 Materials and Methods
2.1 Fungal endophytes
Five fungal endophytes were isolated from the roots of T. wallichiana that were collected from Jageshwar area (2935–2939 N and 7959–7953E), district Almora in Uttarakhand, India. The endophytic fungi, each designated with a code number, were preserved at 4C in a refrigerator. The identification of fungal isolates was based on the polyphasic approach, including phenotypic and molecular characters. Details of isolation, characterization, and identification of these fungal endophytes have been published in our previous study (Adhikari and Pandey, 2019). Fresh cultures were raised for the experimental work.
2.2 Extract Preparation for Paclitaxel Quantification
Extract was prepared by following the previous reported method by Li et al. (2017). Following subculturing of endophytic fungi on Potato Dextrose (PD) agar plate, it was transferred to S-7 fermentation medium and cultured at 25C with centrifugation at 38g. Following 12 days incubation period, the cultures were filtered through sterile filter paper (Whatman size 41) and S7 broth was extracted with an equal volume mixture of ethyl acetate and methanol (v:v 1:1) for 3 h. The organic phase was extracted and collected by a separating funnel. The organic solvents were evaporated under reduced pressure using a vacuum evaporator (Narang Scientific Works, New-Delhi, Model-257) at 45C. The samples were then dissolved in chloroform.
2.3 Screening of Endophytes for Paclitaxel Production
Qualitative and quantitative analysis of paclitaxel was carried out following the previously reported procedure by Nadeem et al. (2019). 20 L of extract from endophytic fungi was injected in HPLC (High performance liquid chromatography) injector (Shimadzu LC solution) equipped with a reverse phase column (C18, 5 m, 2504,6 mm). Methanol: acetonitrile: water in the ratio of (23:20:57 v/v/v) was taken as a mobile phase, at a flow rate of 0.800 mL/min in isocratic mode. Post run analysis of paclitaxel was done at 227 nm wavelength using Photodiode array (PDA) detector. Quantification was done based on standard curve prepared by injecting known concentration of paclitaxel. LOD (limit of detection) and LOQ (limit of quantification) was calculated by standard calibration curve values in the MS excel, by using the equation given NATA (2013) and Sengul (2016). All the fungal endophytes extracts were run in triplicate manner.
2.4 PCR Amplification-based Screening of Potential Paclitaxel Producing Fungi
Endophytic fungal cultures were grown individually in S7 medium at 25C for 74–120 h. Genomic DNA were isolated from the collected mycelia using previously described method Saghai-Maroof et al. (1984). The genes coding for 10-deacetylbaccatin III-10-O-acetyl transferase (DBAT) and C-13 phenylpropanoid side chain- CoA acyltransferase (BAPT) was used as molecular markers to screen paclitaxel producing endophytic fungi. Primer DBAT-F (5-GGGAGGGTGCTCTGTTTG-3), DBAT-R (5GTTACCTGAACCACCAGAGG-3) and BAPT-F (5-CCTCTCTCCGCCATTGACAA-3), BAPT R(5TCGCCATCTCTGCCATA CTT-3) were designed and synthesized following Roopa et al. (2015).
A standard PCR reaction of 25 L consisting 4 L genomic DNA, 1.2 L of forward and reverse primer (10 M), 0.2 L taq polymerase (2U), 1 L of dNTP, 2.5 L of 10X taq buffer (20 mM), 2.5 L of MgCl (4 mM) and 13.6 L of nuclease free water was performed. For DBAT PCR reaction, initially denaturation at 95C (6 min) followed by 35 amplification cycle at 94C (30 s) was done. Annealing was carried out at 50C for 50 s and final extension at 72C for 10 min. Similarly, for BAPT PCR reaction initially denaturation at 95C (6 min) followed by 35 amplification cycle at 94C (50 s) was done. Annealing was carried out at 55C for 55 s and final extension at 72C for 10 min using thermocycler (Bio Rad, USA). The amplified DNA fragment was analysed using agarose gel electrophoresis. A 100 bp size marker was used as a reference (Genaxy, India).
2.5 Optimization of Physico-chemical Parameters for Paclitaxel Production
Effects of different physical and chemical parameters was analyzed for paclitaxel production, initially a range of temperature (5–35C), pH (4 to 11), and incubation time (5–25 days) were tested. Subsequently, influence of protein concentration (peptone), carbon supplements (glucose, fructose, cellulose, starch, maltose, and sucrose) and nitrogen supplements (sodium nitrate, ammonium chloride, ammonium ferrous sulphate and potassium nitrate) on paclitaxel production was determined. A disk of freshly grown fungal culture was inoculated in S7 medium. For paclitaxel quantification HPLC procedure of section 2.4 and 2.5 was followed. The value for each sample was calculated as means of replicates standard error.
2.6 Statistical Analysis
All the experiments were performed in triplicate and data of all the experiments are expressed as the means standard errors (SE). Duncan multiple range test (DMRT) was used for finding their level of significance and for homogenize grouping using SPSS version 20. Statistical significance was determined using ANOVA; p value 0.05 was considered as significant.
3 Results
3.1 Quantitative Estimation for Paclitaxel Production
Initially, the five endophytic fungi, isolated from the roots of T. wallichiana, were investigated for the production of paclitaxel. Detailed information about these T. wallichiana associated fungal endophytes has been described in Adhikari and Pandey (2018; 2019).
The calibration curve for paclitaxel standard showed good linearity with high reproducibility and accuracy at all the tested concentrations (2.15 to 5.45 mg/L). Regression analysis of the calibration curve data points showed an excellent correlation coefficient (R) of 0.9909. The linear regression equation for standard paclitaxel was y 12677x-35449, where x is the concentration of standard and y is the total peak area. The equation thus generated from the curve by the external standard method was used to calculate the amount of paclitaxel present in crude extracts of endophytes. LOD and LOQ was 2.82 g/mL and 7.02 g/mL respectively.
HPLC analysis revealed that out of five endophytes, two endophytes viz. GBPITWR_F1 (Penicillium sp.) and GBPITWR_F5 (Aspergillus sp.) possessed paclitaxel producing potential. With the adopted protocol, in extracts of these two isolates namely GBPITWR_F1 and GBPITWR_F5, paclitaxel peaks were eluted at retention time identical to standard paclitaxel i.e., at 14.15 0.67 min. GBPITWR_F1 and GBPITWR_F5 produced 3.64 0.57 and 5.45 0.98 mg pt/L paclitaxel, respectively.
3.2 PCR Amplification of Paclitaxel Producing Gene
To examine whether GBPITWR_F1 and GBPITWR_F5 have biosynthetic machinery for paclitaxel production, PCR amplification of two genes viz. BPAT and DBAT (main genes responsible for paclitaxel biosynthesis) were carried out. Both the endophytic fungi were found to be positive for DBAT and BAPT genes. For BAPT gene bands of approximately 280 bp and 350 bp, and for DBAT gene bands of approximate 400 bp and 450 bp were visualized for GBPITWR_F1 and GBPITWR_F5 respectively.
3.3 Physico-chemical Optimization of Endophytic Isolates to Maximize Paclitaxel Production
The present study attempted to enhance paclitaxel yield from both the paclitaxel producing endophytes GBPITWR_F1 and GBPITWR_F5 by optimizing physico-chemical conditions. Different parameters like temperature (5–35C), pH (4 to 11), and incubation time (5–25 days) and medium constituents (0.5 to 3 g/L), viz. salt concentration, carbon and nitrogen sources were optimized.
3.3.1 Physical parameters
Influence of incubation time
In both the endophytic fungi, optimum paclitaxel production was observed at different days of incubation (5 to 25 days). GBPITWR_F1 produced maximum paclitaxel i.e., 2.26 0.57 mg pt/L on 20th days, whereas GBPITWR_F5 produced maximum paclitaxel (3.12 0.67 mg pt/L) on 10th day of incubation. In GBPITWR_F5, paclitaxel production start decreasing after 10th day of incubation while in case of GBPITWR_F1 paclitaxel quantity started decreasing after 20th day of incubation. In contrast, biomass production was found to be maximum at 25th day for both GBPITWR_F1 (6.54 g) and GBPITWR_F5 (7.96 g). Based on these results of paclitaxel production at different incubation times, the effects of other parameters were estimated at 20th and 10th days of incubation for GBPITWR_F1 and GBPITWR_F5, respectively.
Effect of temperature and pH
In the present study, temperature (5–35C) and pH (0.5–8) have influenced the production of paclitaxel. For GBPITWR F1, 25C temperature (3.45 0.64 mg pt/L) and 3.5 pH (3.21 0.83 mg pt/L), and for GBPITWR_F5, 15C temperature (3.87 0.82 mg pt/L) and pH 5 (3.54 0.79 mg pt/L) were found suitable for maximum paclitaxel production and good biomass production by the fungal endophytes (Figure 1). Both the endophytic fungi showed higher biomass production at mesophilic temperature range 15–25C.
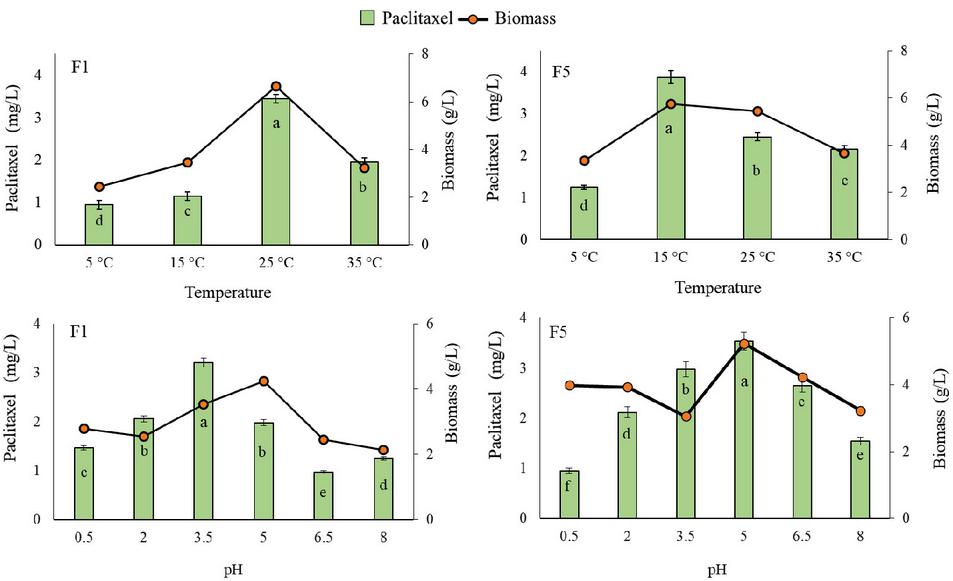
Figure 1 Effect of temperature and pH on taxol production by GBPITWR_F1 and GBPITWR_F5 and biomass erstimation. Results are mean (n 3) SE. Homogenize grouping were done using Duncan test, alphabet a-f denotes their significance (p 0.05).
3.3.2 Chemical parameters
After optimization of physical parameters, chemical constituents were also optimized like protein, carbon and nitrogen source concentration.
Influence of protein concentration
In the present study, peptone was used as a protein source. Different concentrations (ranging from 0.5–5.0 g/L) of peptone were tested to obtain its optimum concentration. 2 g/L peptone was found to be optimum for maximum biomass (5.21 g/L, 8.34 g/L) and paclitaxel concentration (3.05 0.42 mg pt/L, 3.52 0.64 mg pt/L) for both the endophytes GBPITWR_F1 and GBPITWR_F5, respectively. After that, during 3–5 g/L concentration biomass and paclitaxel accumulation started decreasing.
Influence of carbon source
In the present study, different carbon sources including monosaccharides (glucose and fructose), disaccharides (sucrose and maltose) and polysaccharides (starch and cellulose) were tested for fungal biomass growth and paclitaxel production. Results showed that effect of carbon source on paclitaxel production depend on the endophytic fungi. GBPITWR_F1 and GBPITWR_F5 showed maximum paclitaxel and biomass accumulation by using glucose (2.19 0.72 mg pt/L, 6.24 g/L), and maltose (2.98 0.69 mg pt/L, 5.24 g/L) as a carbon source, respectively (Figure 2). After testing the effects of carbon source, concentration of carbon source (ranging from 0.5–3.0 g/L) were also tested for fungal cell biomass growth and paclitaxel production. For GBPITWR_F1 maximum paclitaxel production and biomass i.e., 1.96 0.59 mg pt/L, 5.48 g/L was achieved at 2 g/L of glucose. While for GBPITWR_F5, biomass and paclitaxel accumulation (2.57 0.68 mg pt/L, 6.84 g/L), were higher at 2 g/L of maltose (Figure 2).
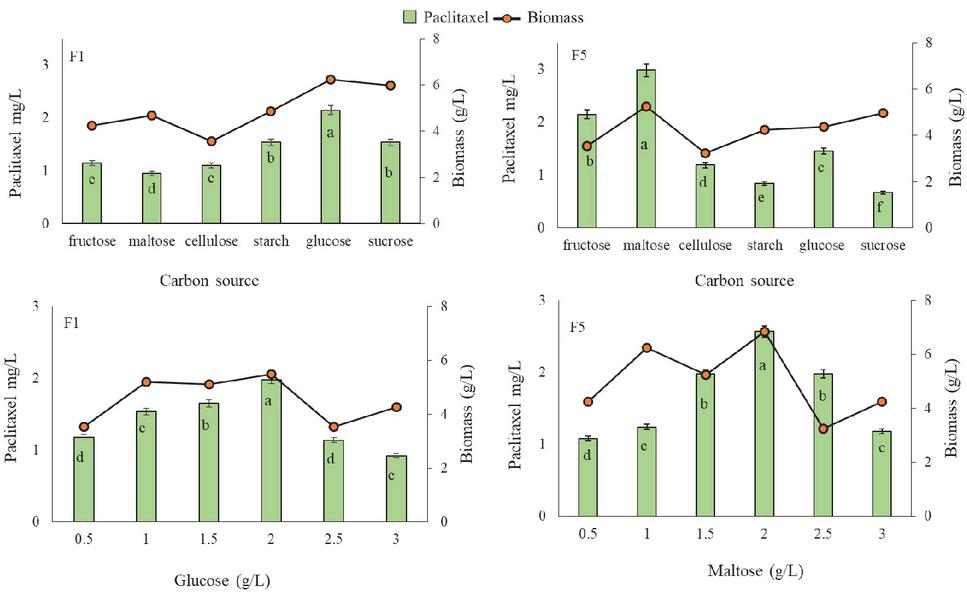
Figure 2 Effect of carbon source on taxol production by GBPITWR_F1 and GBPITWR_F5 and biomass erstimation. Results are mean (n 3) SE. Homogenize grouping were done using Duncan test, alphabet a–f denotes their significance (p 0.05).
Influence of nitrogen source
Effects of four different nitrogen sources i.e., ammonium ferrous sulphate, urea, ammonium chloride, and potassium nitrate were tested on fungal cell biomass growth and paclitaxel production. Accumulation of paclitaxel and biomass of endophytes varied with the nitrogen source used. GBPITWR_F1 and GBPITWR_F5 showed maximum paclitaxel and biomass accumulation by using ammonium ferrous sulphate (2.15 0.26 mg pt/L, 5.87 g/L), and potassium nitrate (2.67 0.39 mg pt/L, 6.84 g/L) as a nitrogen source, respectively (Figure 3). After optimization of nitrogen source, concentration of nitrogen sources (ranging from 0.5-3.5 g/L) were optimized for paclitaxel production. For GBPITWR_F1, paclitaxel production and biomass (2.15 0.64 mg pt/L, 6.45 g/L) was maximum at 1.5 g/L of ammonium ferrous sulphate, whereas for GBPITWR_F5, paclitaxel production and biomass accumulation (2.58 0.62 mg pt/L, 7.54 g/L) both were higher at 1.5 g/L of potassium nitrate (Figure 3).
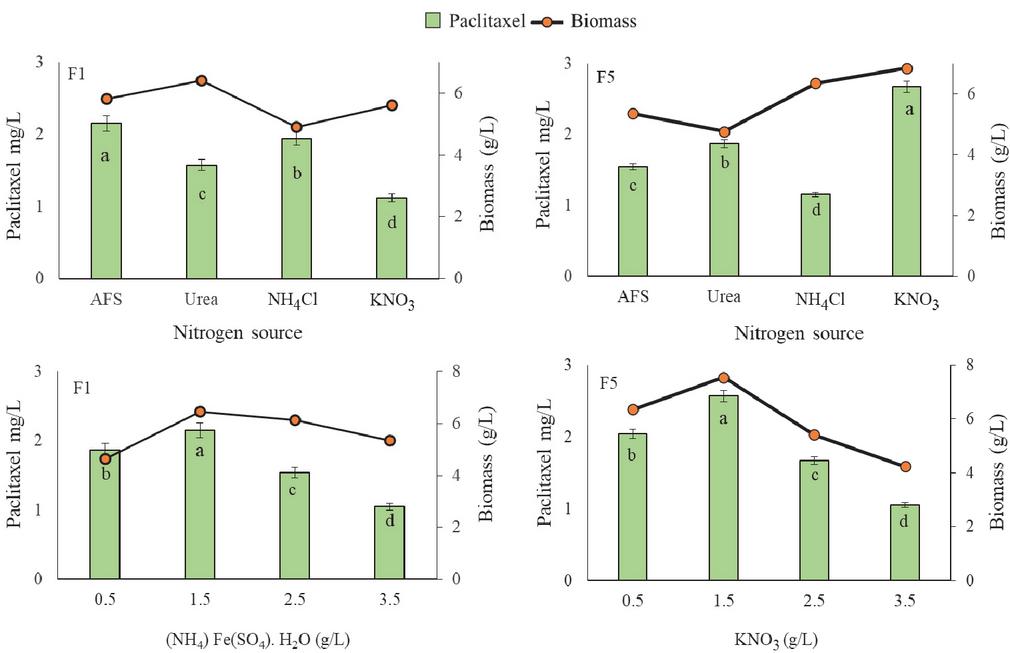
Figure 3 Effect of nitrogen source on taxol production by GBPITWR_F1 and GBPITWR_F5 and biomass erstimation. (NH) Fe(SO). HO (AFS (ammonium ferrous sulphate)), NHCl (ammonium chloride), and KNO (potassium nitrate). Results are mean (n 3) SE. Homogenize grouping were done using Duncan test, alphabet a-d denotes their significance (p 0.05).
3.4 Quantification of Paclitaxel After Media Optimization
After optimization experiments for paclitaxel production by both the endophytes, quantification of paclitaxel production in optimized media viz. Medium 2 (GBPI_TWR F1) and Medium 3 (GBPI_TWR F5) was done. Details of different medium compositions are given in Table 1. Before physico-chemical optimization experiments, GBPI_TWR F5 and GBPI_TWR F1 produced 2.98 0.42 mg/L and 3.15 0.36 mg/L paclitaxel, respectively. After optimization experiments, GBPI_TWR F5 after 10 days of incubation at 15C could produce maximum 5.45 0.98 mg/L paclitaxel on Medium 3 which is S7 liquid medium containing 1 mg/L sodium acetate, 2.5 mg/L zinc sulphate, 1 mg/L copper chloride, 1 mg/L copper sulphate, 3.6 mg/L magnesium sulphate, 2 mg/L ferric chloride, 5 mg/L magnesium chloride, 6.5 mg/L calcium chloride, 2 g/L maltose and 1.5 g/L potassium nitrate, peptone 2 g/L. Final composition of S7 medium after optimization is given in Table 1. The results of physico-chemical optimization experiments illustrated the impact of nutritional manipulation on paclitaxel production by endophytic fungi. Results showed that the type of carbon and nitrogen source influences the accumulation of paclitaxel in fermentation medium. Paclitaxel production in the medium was also influenced by the capacity of endophytic fungi to use the available nutrition sources. For example, in Medium 1 when different combinations of carbon sources were used, biomasses were recorded high with low paclitaxel production. While in Medium 2 and 3, where single carbon and nitrogen sources were used, biomass was not affected but the accumulation of paclitaxel increased in comparison to Medium 1.
Table 1 Medium composition of S7 medium (Media 1), and after optimization Medium 2 and Medium 3
| Medium Composition | Medium 1 | Medium 2 | Medium 3 |
| Sodium acetate | 1 mg/L | 1 mg/L | 1 mg/L |
| Zinc sulphate | 2.5 mg/L | 2.5 mg/L | 2.5 mg/L |
| Copper chloride | 1 mg/L | 1 mg/L | 1 mg/L |
| Copper sulphate | 1 mg/L | 1 mg/L | 1 mg/L |
| Magnesium sulphate | 3.6 mg/L | 3.6 mg/L | 3.6 mg/L |
| Ferric chloride | 2 mg/L | 2 mg/L | 2 mg/L |
| Magnesium chloride | 5 mg/L | 5 mg/L | 5 mg/L |
| Calcium nitrate | 6.5 mg/L | 6.5 mg/L | 6.5 mg/L |
| Glucose | 1 g/L | 2 g/L | Not used |
| Sucrose | 1 g/L | Not used | Not used |
| Fructose | 1 g/L | Not used | Not used |
| Maltose | Not used | Not used | 2 g/L |
| Peptone | 1 g/L | 2 g/L | 2 g/L |
| Ammonium ferrous sulphate | Not used | 1.5 g/L | Not used |
| Potassium nitrate | Not used | Not used | 1.5 g/L |
| Incubation time | 7 days | 20 days | 10 days |
| Temperature | 28C | 25C | 15C |
| pH | 7 | 3.5 | 5 |
| Paclitaxel (mg/L) | |||
| GBPI_TWR F1 | 2.98 0.42 | 3.64 0.53 | 2.15 0.22 |
| GBPI_TWR F2 | 3.15 0.36 | 3.95 0.42 | 5.45 0.48 |
| Biomass (g) | |||
| GBPI_TWR F1 | 4.98 0.15 | 5.26 0.21 | 5.38 0.32 |
| GBPI_TWR F2 | 6.87 0.16 | 5.98 0.25 | 6.87 0.12 |
| Media 1 Composition of S7 media (Li et al., 2017), Medium 2 Composition of S7 media after optimization for GBPITWR_F1, Medium 3 Composition of S7 media after optimization for GBPITWR_F5. | |||
4 Discussion
Bioprospection of endophytic microorganisms is increasingly becoming a major concern for different industries for scale up production of pharmaceutically important bioactive molecules (Kumar et al., 2019). Different Taxus species are known for the accumulation of drug paclitaxel and their endophytes are gaining attention as prospective source for the sustainable and year-round production of paclitaxel without exploiting field grown Taxus plants. Identification of novel paclitaxel producing endophytes will not only facilitate the conservation of Taxus species, but also open a new avenue for sustainable paclitaxel production which is urgently required considering its high demand and price.
HPLC analysis revealed that out of five endophytes, two endophytes viz. GBPITWR_F1 (Penicillium sp.) and GBPITWR_F5 (Aspergillus sp.) possessed paclitaxel producing potential. A recent study reported that fungal genera responsible for paclitaxel production, either isolated from yews or not, are under the phylum Ascomycota (El-Bialy and El-Bastawisy, 2020). Earlier, many paclitaxel producing Aspergillus and Penicillium spp. viz., Aspergillus niger, Aspergillus aculeatinus, Aspergillus fumigatus, Penicillium aurantiogriseum and Penicillium polonicum have been reported from different Taxus species and from other plant species viz., Podocrapus, Corylus avellana and Gingko biloba (Chen et al., 2003; Sun et al., 2008; Yang et al., 2014; Qiao et al., 2017; 2021; Kumar et al., 2019; El-Sayed et al., 2020; Abdel-Fatah et al., 2021). However, so far, only three reports on T. wallichiana endophytes are available (Strobel et al., 1996; Shrestha et al., 2001; Zaiyou et al., 2017). Endophytes viz., Pestalotiopsis microspore, Sporormia minima and Trichothecium sp., have been isolated from the bark and branches of Taxus wallichiana of Nepal (Strobel et al., 1996; Shrestha et al., 2001); and Phoma medicaginis was isolated from the branches of Taxus wallichiana var. mairei of China (Zaiyou et al., 2017).
In contrary to previous studies, in the present study, roots of T. wallichiana plant growing in Western Himalayan Region, were used for endophytes (Aspergillus and Penicillium spp.) isolation and isolated two endophytes were found to produce multi-fold higher concentration of paclitaxel (3.64 0.57 and 5.45 0.98 mg pt/L paclitaxel) in comparison to the previously reported species. Among all earlier studies, Kumar et al. (2019) have reported maximum paclitaxel production i.e., 1590 g/L by Aspergillus fumigatus which was isolated from Taxus sp of Northern Himalayan Region. Higher paclitaxel production recorded in this study from endophytes of T. wallichiana roots growing in Western Himalayan Region might be due to plant part used for endophyte isolation and geographical location where the host plant is growing.
To examine whether GBPITWR_F1 and GBPITWR_F5 have biosynthetic machinery for paclitaxel production, PCR amplification of two genes viz. BPAT and DBAT (main genes responsible for paclitaxel biosynthesis) were carried out. DBAT catalyzes the formation of baccatin III, the immediate diterpenoid precursor of paclitaxel (Walker and Croteau, 2000). Whereas, BAPT catalyzes the selective 13-O-acylation of baccatin III with b-phenylalanoyl- CoA as the acyl donor to form N-debenzoyl-20-deoxypaclitaxel, means it catalyses the attachment of the biologically important paclitaxel side chain precursor (Roopa et al., 2015). Presence of DBAT and BAPT genes in endophytic fungi confirmed that they have independent biosynthetic machinery for paclitaxel production. Similar results were reported in different endophytes such as Aspergillus fumigatus (230 bp BAPT gene), Botryosphaeria rhodina (280 bp DBAT), Trichoderma longibrachiatum (480–720 bp BAPT) and Alternaria sp. (410 bp BAPT) (Roopa et al., 2015; Kumar et al., 2019). Bands of BAPT and DBAT primers in both the endophytic genes, help in assuming that both the endophytes can be used for the production of paclitaxel and paclitaxel related compounds like baccatin III. Alternate possibility for these genes is that they can be used in semisynthetic production of taxanes and paclitaxel.
Optimization of physico-chemical parameters of batch culture is one of the most important strategies in increasing paclitaxel production by endophytic fungi (Naik, 2018). Optimum production of metabolites can be achieved through modification of different biotic and abiotic factors (Somjaipeng et al., 2016). Based on this, the present study attempted to enhance paclitaxel yield from both the paclitaxel producing endophytes GBPITWR_F1 and GBPITWR_F5 by optimizing physico-chemical conditions. Different parameters like temperature, pH, incubation time and medium constituents, viz. salt concentration, carbon and nitrogen sources were optimized. GBPITWR F1, showed maximum paclitaxel production at 25C temperature and GBPITWR_F5, at 15C. There may be many reasons for paclitaxel production in the lower temperature range 5–25C but in present study our view is, as both the endophytic fungi have ability to grow at wide range of temperature 5–35C, detailed study on their growth requirement were reported in earlier study (Adhikari et al., 2018). Moreover, higher paclitaxel production also may be due to the psychrotolerant behaviour of the GBPITWR_F5 fungi, as the average temperature throughout the year remains low in Himalayan mountains.
In GBPITWR_F5, paclitaxel production start decreasing after 10th day of incubation while in case of GBPITWR_F1 paclitaxel quantity started decreasing after 20th day of incubation. On the similar line, maximum paclitaxel production during 21 days of incubation have been reported in Aspergillus terreus isolated from Podocarpus gracilor (El-Sayed et al., 2018) and A. fumigatus isolated from Taxus sp. (Kumar et al., 2019). Based on these results of paclitaxel production at different incubation times, the effects of other parameters were estimated at 20th and 10th days of incubation for GBPITWR_F1 and GBPITWR_F5, respectively. Incubation time plays an important role in production of secondary metabolites in filamentous fungi by providing suitable time to fully proliferate in the medium (Afshari et al., 2015).
In order to achieve maximum production of secondary metabolites, temperature and pH also play crucial role as they are main parameters that affect the physiology and biochemistry of microorganisms and thus affect the metabolite production (Pandey et al., 2018). Different workers reported the effects of temperature and pH on production of secondary metabolites by fungi (Costa et al., 2002; El-Sayed et al., 2018). Likewise, in the present study, temperature (5–35C) and pH (0.5–8) have influenced the production of paclitaxel. Both the endophytic fungi showed higher biomass production at mesophilic temperature range 15–25C. Similarly, endophytic fungi isolated from T. baccata (English yew) i.e., Paraconiothyrium variabile and Epicocum nigrum also produced paclitaxel at pH 5 and 25C (Somjaipeng et al., 2015; 2016).
Secondary metabolites synthesis in the microorganisms can be influenced by manipulating the type and concentration of carbon source of culture media (Sanchez et al., 2010). So, after optimization of physical parameters, chemical constituents were also optimized like protein, carbon and nitrogen source concentration. Protein plays an important role in maintaining nutritional status of the media. Increase and decrease in its concentration may result in slow growth of fungi which directly or indirectly hampers the secondary metabolites production (Andualem and Gessesse, 2013). In the present study, different carbon sources including monosaccharides (glucose and fructose), disaccharides (sucrose and maltose) and polysaccharides (starch and cellulose) were tested for fungal biomass growth and paclitaxel production. Results showed that effect of carbon source on paclitaxel production depend on the endophytic fungi. The high yield of paclitaxel by the endophytic fungi, in the presence of glucose and maltose, could be due to the reason that they stimulate biosynthetic pathways during importing and utilizing process. Second reason may be that the different forms of sugar source may be induced by maltose itself or by glucose produced from maltose during importing or sensing sugar as a carbon source (Choi et al., 2000).
The type of nitrogen source is another key factor affecting cell growth and product formation by microorganisms. In general, the biosynthesis of fungal secondary metabolites is limited by the nitrogen sources (Shu, 2007). It is generally perceived those environmental factors i.e., nitrogen stress, carbon stress, pH conditions, temperature and so on may affect secondary metabolite biosynthesis in fungi. Studies done so far in various fungal systems have revealed the influence of nitrogen source on the regulation of fungal secondary metabolism (Sharma and Jha, 2015). Presence of multifaceted paclitaxel production features in both the endophytic isolates through utilization of different substrate provide an opportunity for detailed study in terms of their storage, subculturing and downstream processing, which can not only release paclitaxel but are also efficient in maintaining the culture for long run commercialization.
5 Conclusion
Present study enumerated that plant microbe-interaction and microbiome studies in high valued medicinal plants are becoming promising and incipient field which helps in identifying the alternative and novel source of microbial drugs. Paclitaxel is one such anticancerous drug which can be isolated from the bark of T. wallichiana; its chemical semi synthesis is not able to satisfy the huge demand. In the present study, two endophytic fungi GBPITWR_F1 (Penicillium sp.) and GBPITWR_F5 (Aspergillus sp.) are determined as potent paclitaxel producer based on the HPLC and presence of molecular biomarker for paclitaxel synthesis. Optimization of physico-chemical properties i.e., temperature, pH, incubation time, carbon and nitrogen source for secondary metabolite production, with particular reference to paclitaxel production, is an important and crucial step. Overall, the present study demonstrates the possibility of biosysnthesis of paclitaxel under proper growth conditions. Both the endophytes could produce good amount of paclitaxel i.e., 3.64 0.57 mg pt/L (GBPI_TWR F1) and 5.45 0.98 mg pt/L (GBPI_TWR F5). Thus, both the endophytes can be considered as alternate sources for production of paclitaxel by pharmaceutical industries. The outcome of this study will be helpful in understanding the biosynthetic mechanism of paclitaxel synthesis in endophytic fungi.
Acknowledgement
Authors are grateful to Director GBPNIHE Almora, Uttarakhand-India, for extending facilities. National Mission on Himalayan Studies (Ministry of Environment, Forest & Climate Change, Govt. of India, New Delhi) is acknowledged for financial support.
References
Abdel-Fatah, S.S., El-Batal, A.I., El-Sherbiny, G.M., Khalaf, M.A., El-Sayed, A.S. (2021). Production, bioprocess optimization and g-irradiation of Penicillium polonicum, as a new paclitaxel producing endophyte from Ginkgo biloba. Biotechnology Report. 30, e00623.
Adhikari, P., Pandey, A. (2019). Phosphate solubilization potential of endophytic fungi isolated from Taxus wallichiana Zucc. roots. Rhizosphere. 9, 2–9.
Adhikari, P., Pandey, A. (2018). Diversity of endophytic fungi associated with Himalayan yew (Taxus wallichiana Zucc.) roots. Proceedings of Himalayan Researchers Consortium. 1, 165–173.
Afshari, M., Shahidi, F., Mortazavi, S.A., Tabatabai, F., Eshaghi, Z. (2015). Investigating the influence of pH, temperature and agitation speed on yellow pigment production by Penicillium aculeatum ATCC 10409. Natural Products Research. 29, 1300–1306.
Andualem, B., Gessesse, A. (2013). Production of microbial medium from defatted brebra (Milletia ferruginea) seed flour to substitute commercial peptone agar. Asian Pacific Journal of Tropical Biomedical. 3, 790–797.
Barbuti, A.M., Chen, Z.S. (2015) Paclitaxel through the ages of anticancer therapy: Exploring its role in chemoresistance and radiation therapy. Cancers. 7, 2360–2371.
Choi, H.K., Kim, S.I., Son, J.S., Hong, S.S., Lee, H.S., Chung, I.S., Lee, H.J. (2000) Intermittent maltose feeding enhances paclitaxel production in suspension culture of Taxus chinensis cells. Biotechnology Letters. 22, 1793–1796.
Costa, E., Teixido, N., Usall, J., Atares, E., Vinas, I. (2002). The effect of nitrogen and carbon sources on growth of the biocontrol agent Pantoea agglomerans strain CPA-2. Letters in Applied Microbiology. 25, 117–120.
Chen, Y.J., Zhang, Z., Wang, Y., Su, Y., Zhang, R. (2003). Screening endophytic fungus to produce paclitaxel from Taxus yunnanensis. Biotechnology. 13, 10–11.
El-Bialy, H.A., El-Bastawisy, H. (2020). Elicitors stimulate paclitaxel production by endophytic fungi isolated from ecologically altered Taxus baccata. Journal of Radiation Research and Applied Sciences. 13, 79–87.
El-Sayed, A.S.A., Safan, S., Mohamed, N.Z., Shaban, L., Ali, G.S., Sitohy, M.Z. (2018) Induction of paclitaxel biosynthesis by Aspergillus terreus, endophyte of Podocarpus gracilior Pilger. upon intimate interaction with the plant endogenous microbes. Process Biochemistry. 71, 31–40.
El-Sayed, R., Ahmed, A.S., Hassan, I.A., Ismail, A.A., El-Din, A.Z.A.K., (2020). Semi-continuous production of the anticancer drug paclitaxel by Aspergillus fumigatus and Alternaria tenuissima immobilized in calcium alginate beads. Bioprocess and Biosystems Engineering. 43, 997–1008.
Kumar, P., Singh, B., Thakur, V., Thakur, A., Thakur, N., Pandey, D., Chand, D. (2019). Hyper production of paclitaxel from Aspergillus fumigatus, an endophytic fungus isolated from Taxus sp. of the northern Himalayan region. Biotechnology Reports. 24, 1–13.
Kusari, S., Singh, S., Jayabaskaran, C. (2014). Rethinking production of paclitaxel (paclitaxel) using endophyte biotechnology. Trends in Biotechnol. 32, 304–311.
Li, D., Fu, D.W., Zhang, Y., Ma, X., Gao, L., Wang, X., Zhau, D., Zhao, K. (2017). Isolation, purification, and identification of paclitaxel and related taxanes from paclitaxel-producing fungus Aspergillus niger subsp. taxi. Journal of Microbiology and Biotechnology. 27, 1379–1385.
Nadeem, M., Rikhari, H.C., Kumar, A., Palni, L.M.S., Nandi, S.K. (2002). Paclitaxel content in the bark of Himalayan Yew in relation to tree age and sex. Phytochemistry. 60, 627–631.
Naik, B.S. (2018). Developments in paclitaxel production through endophytic fungal biotechnology: A review. Oriental Pharmacy and Experimental Medicine. 19, 1–13.
NATA, Technical note 17 (2013). Guidelines for the validation and verification of quantitative and qualitative test methods. National Association of Testing Authorities, Australia. pp. 17–18.
Pandey, N., Jain, R., Pandey, A., Tamta, S. (2018). Optimisation and characterisation of the orange pigment produced by a cold adapted strain of Penicillium sp. (GBPI_P155) isolated from mountain ecosystem. Mycology. 9, 81–92.
Poupat, C., Hook, I., Gueritte, F., Ahond, A., Guenard, D., Adeline, M.T., Wang, X.P., Dempsey, D., Breuillet, S., Potier, P. (2000). Neutral and basic taxoid contents in the needles of Taxus species. Planta Medica. 66, 580–584.
Qiao, W., Ling, F., Yu, L., Huang, Y., Wang, T. (2017). Enhancing paclitaxel production in a novel endophytic fungus, Aspergillus aculeatus Tax-6, isolated from Taxus chinensis var. mairei. Fungal Biology. 121, 1037–1044.
Qiao, W., Tang, T., Ling, F. (2021). Comparative transcriptome analysis of a paclitaxel-producing endophytic fungus, Aspergillus aculeatinus Tax-6, and its mutant strain. Scientific Reports. 10, 10558. https://doi.org/10.1038/s41598-020-67614-1.
Roopa, G., Madhusudhan. M.C., Sunil, K.C.R., Lisa, N., Calvin, R., Poornima, R., Zeinab, N., Kini, K.R., Prakash, H.S., Geetha, N. (2015). Identification of paclitaxel producing endophytic fungi isolated from Salacia oblonga through genomic mining approach. Journal of Genetic Engineering and Biotechnology. 13, 119–127.
Saghai-Maroof, M.A., Soliman, K.M., Jorgensen, R.A., Allard, R.W. (1984). Ribosomal DNA spacer-length polymorphism in barley: Mendelian inheritance, chromosomal location and population dynamics. Proceedings of National Academy of Science USA. 81, 8014–8018.
Sanchez, S., Chavez, A., Ferero, A., Huante, Y.G., Romero, A., Sanchez, M., Richa, D., Sanchez, B., Avalos, M., Trampe, S.G., Sanoja, R.R., Langley, E., Ruiz, B. (2010). Carbon source regulation of antibiotic production. Journal of Antibiotics. 63, 442–459.
Sengul, U. (2016). Comparing determination methods of detection and quantification limit for oflatoxin analysis in hazulnut. Journal of Food Drug and Analysis. 24, 56–62.
Sharma, B., Jha, D.K. (2015). Role of nitrogen sources in regulation of fungal secondary metabolism. In: Gupta VK, Mach RL, Sreenivasaprasad S (eds) Fungal biomolecules: Sources, applications and recent developments, 1st edn. John Wiley & Sons, Ltd., pp. 213–224.
Shreshta, K., Strobel, G.A., Shrivastava, S.P., Gewali, M.B. (2001). Evidence for paclitaxel from three new endophytic fungi of Himalayan Yew of Nepal. Planta Medica. 67, 374–376.
Shu, C.H. (2007). Fungal fermentation for medicinal plants. In: Yang ST (ed) Bioprocessing for value-added products from renewable resources, 1st edn. Elsevier, pp. 447–463.
Somjaipeng, S., Median, A., Kwasna, H., Ordaz-Ortiz, J., Magan, N. (2015). Isolation, identification and ecology of growth and paclitaxel production by an endophytoc strain of Paraconiothyrium variabile from English Yew tree (Taxus baccata). Brazilian Mycological Society. 119, 1022–1031.
Somjaipeng, S., Medina, A., Magan, N. (2016). Environmental stress and elicitors enhance paclitaxel production by endophytic strains of Paraconiothyrium variabile and Epicoccum nigrum. Enzyme and Microbial Technology. 90, 69–75.
Sun, D., Ran, X., Wang, J. (2008). Isolation and identification of a paclitaxel-producing endophytic fungus from Podocarpus. Acta Microbiologica Sinica. 48, 589–595.
Thomas, P., Farjon, A. (2011). Taxus wallichiana. The IUCN Red List of Threatened 2011:e.T46171879A9730085.
Stierle, A., Strobel, G., Stierle, D., 1993. Taxol and taxane production by Taxomyces andreanae, an endophytic fungus of Pacific yew. Sci. 260, 214–216.
Strobel, G.A., Yang, X., Sears, J., Kramer, R., Sidhu, R., Hess, W.M. (1996). Paclitaxel from Pestalotiopsis microspora, an endophytic fungus of Taxus wallichiana. Microbiology. 142, 435–440.
Wani, M.C., Taylor, H.L., Wall, M.E., Coggon, O., McPhail, A.T. (1971). Plant antitumor agents VI. The isolation and structure of paclitaxel, a novel antileukemic and antitumor agent from Taxus brevifolia. Journal of American Chemical Society. 92, 2325–2327.
Walker, K.D., Croteau, R.B. (2000). Molecular cloning of a 10-deacetylbaccatin III-10-O-acetyl transferase cDNA from Taxus and functional expression in Escherichia coli. Proceedings of National Academic Science. 97, 583–587.
Yang, Y., Zhao, H., Barrero, R.A., Zhang, B., Sun, G., Wilson, L.W., Xie, F., Walker, K.D., Parks, J.W., Bruce, R., Guo, G., Chen, L., Zhang, Y., Huang, X., Tang, Q., Liu, H., Bellgard, M.I., Qiu, D., Lai ,J., Hoffman, A. (2014). Genome sequencing and analysis of the paclitaxel-producing endophytic fungus Penicillium aurantiogriseum NRRL 62431. BMC Genomics. 69. https://doi.org/~10.1186/1471-2164-15-69
Zaiyou, J., Li, M., Xiqiao, H. (2017). An endophytic fungus efficiently producing paclitaxel isolated from Taxus wallichiana var. mairei. Medicine. 96, e7406. https://doi.org/10.1097/MD.0000000000007406
Biographies

Priyanka Adhikari is Research Associate at National Institute of Pharmaceutical Education and Research (NIPER), Guwahati-Assam, India. Before Joining NIPER, Dr. Adhikari worked as Junior and Senior Project Fellow in National Mission on Himalayan Studies (NMHS) Program of Ministry of Environment, Forest and Climate Change, Govt. of India at G.B. Pant National Institute of Himalayan Environment, Almora – Uttarakhand, India. Dr. Adhikari has M.Sc. in Microbiology and Ph.D. in Biotechnology from Kumaun University, Nainital-Uttarakhand, India. Her research interests include plant-microbe interactions and characterization, and identification of microbe and plant based bioactive compounds and drug formulation. Dr Adhikari has been recognized for National Award (twice) from MoEF & CC Himalayan Researchers Consortium and National Medicinal Plant Board (NMPB) Govt. of India for her work on microbiological and biochemical aspects of Taxus wallichiana. Later, she received the Young Scientist Travel Grant to present her work in 8th Conference “FEMS 2019” at Glasgow Scotland, United-Kingdom. She has published more than 20 peer reviewed papers in National and International Journals.

Mithilesh Singh is working as Scientist at Centre for Environmental Assessment and Climate Change, G.B. Pant National Institute of Himalayan Environment (GBP-NIHE), Almora, Uttarakhand. She has done post graduation in Botany from Banaras Hindu University and PhD in Biotechnology from IIT Guwahati, Assam. Her field of specialization includes plant biotechnology and bioprospection. She has published 55 research papers, chapters in edited books/proceedings and scientific popular articles having over 520 citation and 12 h index. Dr. Singh has received DBT BioCARe Women Scientist Award in 2014.

Anita Pandey is presently affiliated with the Department of Biotechnology, Graphic Era (Deemed to be University, Dehradun, Uttarakhand, India. Previously, she was Scientist at G.B. Pant National Institute of Himalayan Environment, Almora, India. Dr. Pandey has extensively worked on Bioprospection of Microbial Diversity of Indian Himalayan region (IHR). Her research areas include- Extremophiles, Rhizosphere, Biodegradation, Fermented Foods, and Antimicrobials.
Her scientific contributions have been recognized at State, National, and International levels. She has been recipient of National Bioscientist Award by Department of Biotechnology, Govt. of India and Vishisht Mahila Vaigyanik Sammaan (Eminent Woman Scientist Recognition) by Uttarakhand Science, Education and Research Center, Department of Science and Technology, Govt. of Uttarakhand, India.
Dr. Pandey has hosted CV Raman International Fellowship for African Researchers twice (Morocco and Egypt). A new archeal species (Nantronococcus pandeyae sp. nov.) has been named in her honour for her extensive contribution to microbial diversity of IHR (Current Microbiology, vol. 79, Article number 51, 2022).
Journal of Graphic Era University, Vol. 10_2, 195–216.
doi: 10.13052/jgeu0975-1416.1028
© 2022 River Publishers